The gland

Anterior pituitary cells
The anterior pituitary is made up of a number of cell types that are essential for such physiological processes as growth, development, homeostasis, metabolism and reproduction.
In the anterior pituitary there are 2 main different kind of cells: acidophil and basophil, the names of the cells is due to the affinity of the colorant used in histochemistry (acid or basic colorant).
The term "
acidophil" is used to describe two different types of cells:
- somatotropes (growth hormone ( GH ) producing cells)
- lactotropes (prolactin (PRL) producing cells).
The term "
basophil" is used include the following cell types:
- corticotropes (adrenocorticotropic hormone (ACTH) and related peptides producing cells)
- thyrotropes (thyroid stimulating hormone (TSH) producing cells)
- gonadotropes (luteinizing hormone (LH) and follicle stimulating hormone (FSH) producing cells).
The acid-basic characteristic of the pituitary cells is due to the hormone they contain in granules. From the analysis of the aminoacidic sequence is possible to calculate the protein aminoacids percentage (graph 1, graph 2) and the isoelectric point of the proteins(graph3):

Graph 1 - Aminoacids percentage in acidophil cells.

Graph 2 - Aminoacids percentage in basophil cells.

Graph 3 - Isoelectric point of the proteins in acidophil cells and in basophil cells



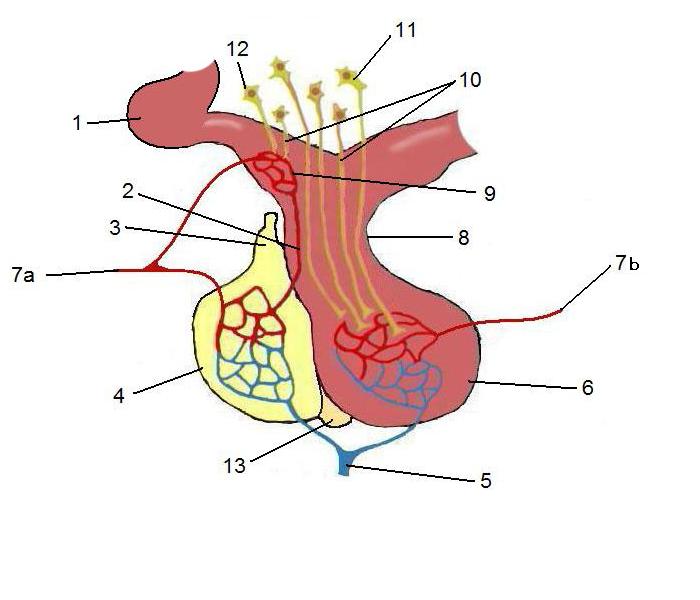
But acidophilic and basophilc cells aren’t only characterized by different pattern of production of hormones, but also by they localization in the anterior pituitary. Acidophilic cells are most adboundant in the antero-lateral area, the basophilic cells in the posterior-central area.
Even after differentiation, pituitary cells continue to undergo mitosis and this process could be augmented under certain conditions in adulthood.
There is increasing evidence indicating that some anterior pituitary cells are multifunctional in nature and could exhibit mixed phenotypes. Interestingly, multifunctional cells are found to be more abundant in females than in males, indicating that the hormonal changes associated with the cyclical events in females may promote transdifferentiation.
Pituitary cell differentiation and growth are dependent on many locally produced and released molecules acting in autocrine or paracrine manners. These include nitric oxide, ATP, acetylcholine and peptide factors such as endothelins, activins, neurotropins, leukemia inhibitory factor and insulin-like growth factors.
Pituitary Development
The cells destined to become the endocrine pituitary gland are located in the midline of the anterior neural ridge. The neural ridge forms a pocket (Rathke's pouch ) that comes into contact with, and invaginates into, the ventral diencephalon. Together they will form the pituitary, the anterior (endocrine) pituitary originating from Rathke's pouch, and the posterior (neural) pituitary originating from the ventral diencephalon.
The three major stages in the formation of the anterior pituitary gland are: (1) extrinsic signals that cause cell proliferation and determination as pituitary; (2) intrinsic signaling gradients within Rathke's pouch activate a core group of transcription factors; and (3) commitment of cells to particular lineages through combinatorial associations of transcription factors.
Stage 1: Initial proliferation and induction.
The first stage of anterior pituitary growth and differentiation involves extrinsic paracrine signals from the diencephalon (dorsally) and from the pharyngeal (oral) ectoderm ventrolaterally (Figure 1). The diencephalon cells produce BMPs, Wnt5a, and FGF10. The oral ectoderm produces Sonic hedgehog. FGFs and sonic hedgehog are critically important at this stage. BMP4 is required for the cell division of Rathke's pouch. Sonic hedgehog is expressed throughout the oral ectoderm, except in the region that is destined to become Rathke's pouch. This combination of signals, from top and bottom, cause the Rathke's pouch cells to express the transcription factor LHX3, a critical factor for specifying these cells as the precursors of the endocrine cell types.

Figure 1 - Initial proliferation and induction.
Stage 2: Intrinsic signaling and specification.
The patterning of Rathke's pouch is established by intrinsic gradients of paracrine factors within the ectoderm, itself, and from the condensing mesenchyme associated with the pouch. First a ventral-to-dorsal gradient of
BMP2 and Indian hedgehog is established within the ventral ectoderm of the pouch (BMP2) and in the condensing ventral mesenchyme (BMP2,
Ihh). In the opposite direction is a dorsal-to-ventral gradient of
FGFs. These gradients cause overlapping sets of transcription factors to be synthesized in different populations of cells, according to their positions along the dorsal-ventral axis (Figure 2). Many of these transcription factors are required to regulate the expansion of particular sets of multipotent precursor cells.

Figure 2 - Intrinsic signaling and specification.
Stage 3. Cell commitment.
The information in the transient gradients of paracrine factors becomes stabilized in the discrete pattern of transcription factor synthesis (Figure 3).

Figure 3 - Cell commitment.
Figure 4 shows the lineages of the anterior pituitary gland with respect to their transcription factors. At early stages, these cells are thought to be multipotent, but their fate becomes progressively limited by the sets of transcription factors contained within the nucleus. LHhx3-positive cells with no Prop-1 transcription factor become the rostral tip thyrocytes. Those cells acquiring transcription factor Tbx19 become the ventral corticotropes and the melanotropes, while the majority of pituitary cell types are derived from the cells that acquire the Prop-1 transcription factor.

Figure 4 - Cell lineages of the anterior pituitary gland.
Of the Prop-1 positive cells, the ventral cells synthesize the GATA-2 transcription factor and make gonadotropes and thyrotropes. The dorsal cells of the Prop-1 lineage synthesize the PIT-1 transcription factor and generate the somatotropes, lactotropes, and also the thyrotropes. At high levels, PIT-1 inhibits the transcription of Gata-2, and GATA-2 inhibits the transcription of Pit-1. At lower levels of these factors, both can be synthesized. When both Gata-2 and Pit-1 are activated, the PIT-1 protein can bind to its binding sites adjacent to Gata-2 sites and will activate the thyrotrope genes. Moreover, the PIT-1 protein will bind directly to the GATA-2 protein (with no DNA binding) and block its ability to activate the gonadotroph-specific genes.
The regulation of which Pit-1-positive cells become somatotrophs and which become lactotropes is very complicated. Multiple PIT-1 binding sites are involved in the genes producing growth hormone and prolactin, and the spacing between the PIT-1 binding sites of these genes may be critical in determining which hormone is made. It is possible that both cell types have the potential to make both hormones, but that cell-type-specific repressors are responsible for determining which genes become active. Moreover, the repression may be controlled by timing of whether activators or repressors bind first.