DEFINITION
Hepatitis C virus is a small (55-65 nm) enveloped RNA virus belonging to the Flaviviridae family.
HCV is the cause of Hepatitis C in human.
HCV is a leading cause of chronic liver desease, including cirrhosis and hepatocellular carcinoma.
GENOMIC ORGANIZATION
The HCV genome consists of a single-stranded positive-sense RNA of 9.6 kb. The HCV RNA encodes a single polyprotein that is cleaved into three structural proteins (core, E1 and E2) and seven non structural proteins (p7, NS2, NS3, NS4A, NS4B, NS5A, NS5B).

Genomic organization of HCV
New insights into structure and replication of the hepatitis C virus and clinical implications.2010
LIFE CYCLE
Hepatitis C viral life cycle.2007
1. Attachment and entry: in order to enter the host cell, the virus must first bind to a receptor on the cell surface. The specific interaction between host cell receptors (CD81, SRBI, claudin-1) and viral attachment proteins on the surface of the virion determines tissue tropism and host range. HCV can enter the cell through clathrin-dependent endocitosis.
2. Uncoating: HCV loses the envelope and the genome is released in the cytoplasm.
3. Translation: mRNA is translated via a cap-indipendent IRES-mediated process. The translation product is a large precursor polyprotein that is processed by cellular and viral proteases into mature structural and non structural proteins.
4. RNA replication: it starts with the synthesis of a complementary negative-strand RNA using the genome as a template, after which genomic positive-strand RNA is produced from a negative-strand RNA template; both steps are catalyzed by the NS5B RNA-dependent RNA polymerase.
5. Viral assembly: Little is known about the assembly of HCV. The mature virion consists of a nucleocapsid and outer envelope composed of a lipid membrane and envelope proteins.
6. Maturation
7. Release: HCV is released from the cell through the secretory pathway.

The HCV life cycle
EPIDEMIOLOGY
According to the World Health Organization there are 170 million people infected with HCV, corresponding to 3% of the world's total population. HCV infection considered as major health threat and its distribution among the world differs widely, with the highest incidence in Asia and Africa.

The distribution of HCV infection
There are several genotypes of HCV: genotype 1 is more prevalent in Europe and America, genotype 4 in Africa and genotype 6 in Asia.
Epidemiology and natural history of HCV infection.2013
Most of the millions who are chronically infected with HCV are now in their fourth or fifth decade of life. As they move into their 60s and 70s, these individuals constitute an "age wave" of asymptomatic HCV infection that is headed toward clinical disease.
Rates of HCV infection are also generally higher among men and African Americans.
This phenomenon of an aging generational cohort of HCV-infected patients is also a result of the rapidly declining incidence of new HCV infections over the past 25 years. These positive trends can be attributed to improved blood donor screening and the related reduction in transfusion-associated cases, but even more so to safer needle practices among injection drug users due to concern about infection with human immunodeficiency virus (HIV).

Chronic hepatitis C: an age wave of disease burden.2005
TRASMISSION MODE
• Trasfusion
• Drug addiction
• Nosocomial
• Sexual
• Upright/perinatal
DIAGNOSIS
Laboratory test
1. ELISA to search anti-HCV antibodies: Detection of anti-HCV antibodies in plasma or serum is based on identification of IgG antibodies against several HCV antigens. There are currently no assays for anti-HCV IgM antibodies.
2. Confirmation of positive samples with immunosorbent assay on strip
3. Amplification by RT-nested PCR and sequence analysis of NS5B
The multiple challenges in HCV diagnosis can be divided into the following three groups: false negative test results during a long seronegative window period; significant and varying levels of false positive test results; differentiation between current/chronic infection and cleared/resolved infection. In addition, differentiation between recent and nonrecent infection would benefit both the patient (choice of treatment regime) and society (identification of sources of new infections).
New Tools in HCV Diagnosis, in Light of the Enhanced Awareness and the New Drugs for Treatment.2013
Biopsy
Liver biopsies are used to determine the degree of damage to the liver, however, the procedure involves risks and in 1 to 5% of cases requires hospitalization. Changes observed are typical lymphocytic infiltration within the liver parenchyma, the presence of lymphoid follicles localized in the portal triad and the alteration of the bile ducts.
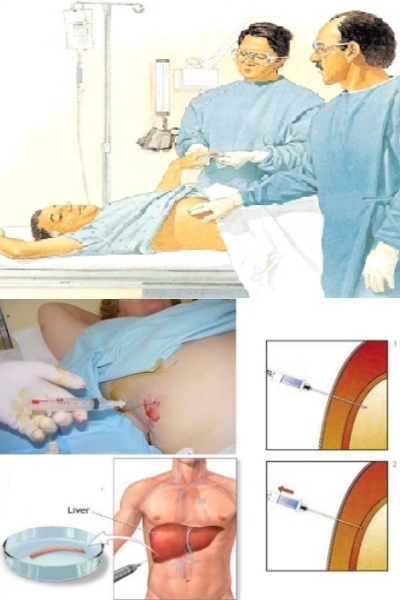
Liver biopsy
Recently, several non-invasive tests have been developed:
• Fibroscan
• FibroMeter
• Fibrotest

Schematic reliability of diagnostic means
Comparison of accuracy of fibrosis degree classifications by liver biopsy and non-invasive tests in chronic hepatitis C.2011
PATHOGENESIS
Frequently the primary infection is asymptomatic, but it causes a chronic persistent infection. It often progresses to active hepatitis within 10-15 years and to cirrhoses and liver failure after 20 years. HCV promotes the development of hepatocellular carcinoma after 30 years in more than 5% of chronic patients.
The ability of HCV to achieve cell death and to remain associated with the host cell allows the establishment of a persistent infection and it causes a liver damage in the long term. The cell-mediated immunopathology is primarily responsible for the tissue damage. The continuous repair mechanism of the liver and the induction of cell growth that occur in chronic HCV are predisponing factors in the development of carcinoma.
HCV escape
HCV has adapted multiple mechanisms to escape host immunity. The low-fidelity HCV RNA polymerase NS5B introduces point mutations to generate a genetic diversity within the quasispecies in an infected host. Many of the viable variants cluster in high-variability regions (HVR1) of the glycoprotein sequences, which can contribute to differential binding and usage of HDL and SR-BI of different variants, providing flexibility for adaptation against host humoral immunity. Glycosylation sites conceal important functional domains of E2 by forming a glycan shield that reduces the viral immunogenicity and the access of the epitopes to nAbs. Additionally, the action of nAbs may also be disturbed by the presence of interfering antibodies. Another way of escaping neutralizing antibodies is by avoiding the circulation altogether, by dissemination via cell-to-cell transmission.

Strategies of HCV evasion
Neutralizing Antibodies and Pathogenesis of Hepatitis C Virus Infection.2012
Interaction HCV-mithocondria
HCV proteins appear to accumulate at points of contact between mitochondria and the endoplasmatic reticulum. This led to the proposal that certain HCV proteins migrate to mitochondria by lateral diffusion from the ER via a transient fusion of the membraneous sub-compartments.
Mitochondria form a dynamic reticulum intertwined with the ER. High-resolution imaging studies proved that the two membranous sub-cellular compartments physically contact each other in many points. These contact sites are not simply a consequence of contiguity but constitute specialized functional areas. Molecular determinants of these contact sites are beginning to be disclosed. The main role of these intracellular “synapses” would be to control the Ca2+ flux between the two organelles. The mitochondrial Ca2+ uniport is limited by its low km for the substrate whereby a relatively high concentration of cytosolic Ca2+ would be required to achieve an efficient inward current. The mitochondria-ER contact site provides a microcompartment where the concentration of Ca2+, released by the ER, can reach localized high concentrations thereby enabling the functioning of the mitochondrial uniporter.
By this way, the ER may communicate with mitochondria via Ca2+ signaling without rising the bulk cytosolic Ca2+ level over a physiological threshold.
Targeting mitochondria in the infection strategy of the hepatitis C virus.2013
HCV-related mitochondrial dysfunctions
It is proposed that the bulk of HCV proteins accumulate at the ER and ER-derived modified membranes, a portion that specifically localizes to ER–mitochondria contact sites is responsible for the HCV-linked mitochondrial alterations. The priming event is the release of Ca2+ from ER stores by opening of the Ca2+ channels. The mechanism is not fully elucidated but a direct interaction or an ER stress-mediated indirect effect can both be considered and are not mutually exclusive.
It has been recently reported that increased ER–mitochondrial coupling is observed during the ER stress. It was shown that components of the ER-UPR increase the expression of a truncated variant of SERCA1. This localizes to and increases the ER–mitochondria contact sites determining progressive Ca2+ leak from the ER and Ca2+ accumulation in the mitochondria. In the early phase of the ER stress a spatial re-organization of the interacting ER and mitochondrial network occurre and this correlate with an increase in ATP levels, oxygen consumption, reductive power and increased mitochondrial Ca2+ uptake.
A further point to be considered is that ROS production and oxidative stress. ROS generation can be upstream or downstream UPR targets and may display a UPR-specific plus a nonspecific component.
The activated outward flux of Ca2+ from the ER enhances its local concentration at ER–mitochondria “synapses” without increasing the bulk cytoplasmic Ca2+ concentration. The entry of Ca2+ into mitochondria is mediated by the voltage-dependent anion channel (VDAC)/porin of the OMM and by the mt�� -driven Ca2+ uniporter of the IMM.
Direct interaction of an HCV protein with these mitochondrial transport systems cannot be excluded. Inter-membrane transfer of proteins from the ER to the OMM is a possible mechanism accounting for the reported localization of HCV proteins on mitochondria. Core, NS3-4A and NS5A might be possible specific effectors in case of a direct interaction with mitochondrial components. The main mitochondrial exit route of Ca2+ is mediated by the Na+/Ca2+ exchanger and by transient opening of the MPTP. Re-uptake of Ca2+ by the ER is dependent on the Ca ATPase. Enhanced recycling of Ca2+ is expected to decrease the ��H+ and thereby the efficiency of ATP synthesis.
High levels of mitochondrial Ca2+ may cause changes in the redox balance by a number of mechanisms. Ca2+ can detach cytochrome c from the inner membrane slowing down the flux of electrons between cytochrome c reductase and the cytochrome c oxidase.
In addition, Ca2+ activates the mitochondrial isoform of a nitric oxide synthase, thereby releasing NO• which is an inhibitor of cytochrome c oxidase. The synergistic combination of these two events causes accumulation of reducing equivalents upstream of cytochrome c oxidase.
The respiratory chain complex which under these conditions predominantly releases ROS is the NADH-dehydrogenase (complex I). Redox active centers and/or nearby residues proved to be targets of superoxide which is generated by the complex I itself therefore establishing a kind of self-inhibition. Moreover, NO and Ca2+ can inhibit RC complex I, enhancing ROS production by this complex. In order to counterbalance this increase of ROS production the intramitochondrial redox buffer (mainly GSH) may become progressively depleted. Rereduction of oxidised glutathione (GSSG) by glutathione reductase requires NADPH. The reduction observed in mitochondria of HCVinfected cells would limit the trans-hydrogenase activity, leading in turn to accumulation of GSSG.
ROS and Ca2+ are known activators of the MPTP. Opening of the pore enables flush out of low molecular weights compounds comprising NAD+/NADH and GSH thereby contributing to further impairment of OXPHOS and of antioxidant buffering.
Accumulation of ROS proved to affect the activity of both the ER Ca2+ channels and mitochondrial calcium channel/transporters.

Overview of alterations in mitochondria linked to HCV infection
Low levels of ROS and mt-Ca2+-dependent stress activates a pro-survival and proliferative adaptive response by redox signaling. The flickering balance between the PTP closed/open
configuration is set to a level that cause collapse of the RC-mediated protonmotive force with consequent impairment of ATP synthesis by the ATP synthase. This forces the infected cell to shift its energy-supplying metabolism toward glycolysis by activation of the transcription factor HIF-1. Collapse of the ��H+ is a trigger for selective removal of damaged mitochondria by the autophagic machinery which recently has been shown to be required for HCV replication. Such a prosurvival setting in the host cell enables HCV maintenance however if additional hits accumulate over the time this may result in clonogenic expansion leading to hepatocarcinoma.
Intermediate levels of ROS and mt-Ca2+ enhances the closed to open transition of the PTP causing, among the other, depletion of low-molecular weight metabolites (NAD+, carnitine, coenzyme A) needed to import long chain acyl-CoA for �-oxidation. Therefore AcCoA accumulate into the cytoplasm and are converted in triglycerides. Other factors, like activation of HIF-1 and of the sterol regulatory element-binding protein (SREBP-1c) may contribute to enhanced de novo lipogenesis. All together this may account for the steatotic development which features the HCV-infected hepatocyte.
High intramitochondrial concentrations of Ca2+ and ROS induces permanent opening of the PTP causing osmotic swelling and rupture of the OMM. The consequent release of cytochrome c and other pro-apoptotic factors triggers the caspase-cascade. Depending on the intracellular ATP level, this would lead to apoptosis or necrosis activating, in the last case, tissue fibrosis.

Consequences of the HCV-induced deregulation of the mitochondrial Ca2+ and ROS homeostasis
Hepatitis C Virus-Induced Mitochondrial Dysfunctions.2013
HCV CONSEQUENCES
• Fibrosis and cirrhosis
• Hepatocellular carcinoma
• Cholangiocarcinoma
• Hepatic decompensation
• Steatosis
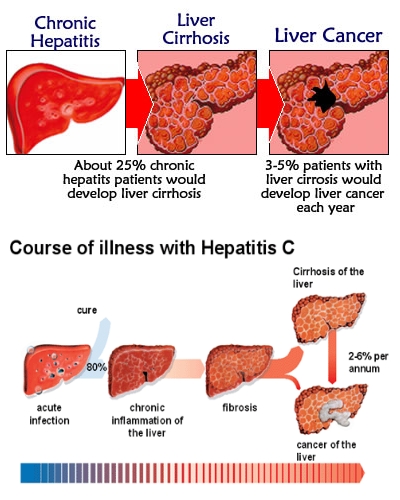
Progression to cirrhosis and hepatocellular carcinoma
Prevalence and challenges of liver diseases in patients with chronic hepatitis C virus infection.2010
THERAPY
The treatment of reference is a combination of pegylated interferon alfa and ribavirin, to be taken for a period of 24 or 48 weeks, depending on the genotype of HCV. It is observed that this therapy leads to improvements in 50-60% of cases. Genotypes 1 and 4 are less sensitive to interferon. In 2011 it has been approved two new antiviral drugs, Baceprevir and Teloprevir, flanking interferon and ribavirin against genotypes more difficult to treat. The treatment that takes place during the first six months is more effective than when hepatitis C becames chronic.
This therapy involves numerous side effects such as flu-like symptoms, alteration of hematological parameters, gastrointestinal and dermatological disorders. The interruption of treatment because of side effect occurs in 10-20% of cases.
Researchers are studying new targets to inhibit the replication cycle of HCV through inhibition of viral proteins.
The molecular and structural basis of advanced antiviral therapy for hepatitis C virus infection.2013
They are also studyng a vaccine against HCV and this is very difficult because HCV has endless mimetic capacity and it is able to escape the immune system thanks to the continuoius development of mutants.
Ever closer to a prophylactic vaccine for HCV.2013.