cGMP play a key role in the visual system by regulating the recovery phase of visual excitation and adaptation to background light. The photoreceptor cells involved in vision are the rods and cones. These cells contain a chromophore (11-cis-retinal, the aldehyde of Vitamin A1 and light-absorbing portion) bound to a cell membrane protein, opsin.
Photransduction-Wikipedia
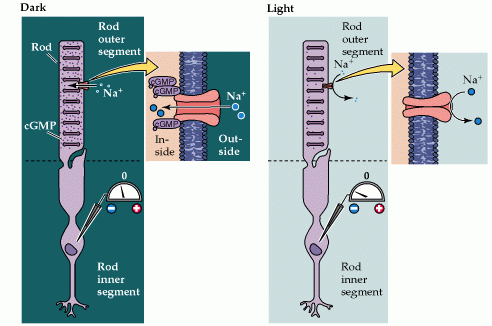
In the Dark
Photoreceptor cells are strange cells because they are depolarized in the dark, i.e. light hyperpolarizes and switches off these cells, and it is this ‘switching off’ that activates the next cell and sends an excitatory signal down the neural pathway.
In the dark, cGMP levels are high and keep cGMP-gated sodium channels open allowing a steady inward current, called the dark current. This dark current keeps the cell depolarised at about -40 mV.
The depolarization of the cell membrane opens voltage-gated calcium channels. An increased intracellular concentration of Ca2+ causes vesicles containing special chemicals, called neurotransmitters, to merge with the cell membrane, therefore releasing the neurotransmitter into the synaptic cleft, an area between the end of one cell and the beginning of another neuron. The neurotransmitter released is glutamate, an excitatory neurotransmitter.

Photoreceptors cell
In the light
Light stimulation of rhodopsin in the receptor disks leads to the activation of a G-protein (transducin).The GTP-bound alpha subunit of transducin activates a phosphodiesterase (PDE).The activated phosphodiesterase hydrolyzes cGMP into GMP, reducing its concentration in the outer segment and leading to the closure of sodium channels in the outer segment membrane.
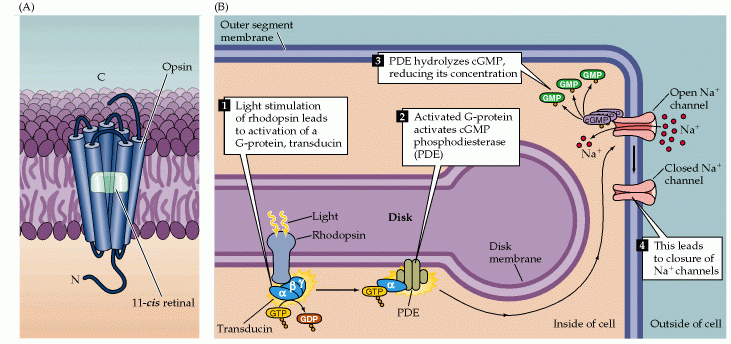
Deactivation of the phototransduction cascade
GTPase Activating Protein (GAP) interacts with the alpha subunit of transducin, and causes it to hydrolyse its bound GTP to GDP, and thus halts the action of phosphodiesterase, stopping the transformation of cGMP to GMP.
Finally, Metarhodopsin II is deactivated. Recoverin, another calcium binding protein, is normally bound to Rhodopsin Kinase when calcium is present. When the calcium levels fall during phototransduction, the calcium dissociates from recoverin, and rhodopsin kinase is released, when it proceeds to phosphorylate metarhodopsin II, which decreases its affinity for transducin. Finally, arrestin, another protein, binds the phosphorylated metarhodopsin II, completely deactivating it. Thus, finally, phototransduction is deactivated, and the dark current and glutamate release is restored. It is this pathway, where Metarhodopsin II is phosphorylated and bound to arrestin and thus deactivated, which is thought to be responsible for the S2 component of dark adaptation. The S2 component represents a linear section of the dark adaptation function present at the beginning of dark adaptation for all bleaching intensities.
Domek-Łopacińska K. et al.,Cyclic GMP metabolism and its role in brain physiology., J Physiol Pharmacol. 2005