Beta-glucan
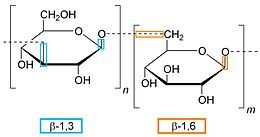
β-Glucans (beta-glucans) are polysaccharides of D-glucose monomers linked by β-glycosidic bonds. β-glucans are a diverse group of molecules that can vary with respect to molecular mass, solubility, viscosity, and three-dimensional configuration. Some forms of beta glucans are useful in human nutrition as texturing agents and as soluble fibersupplements, but can be problematic in the process of brewing.
Glucose molecule, showing carbon numbering notation and β orientation.
The most active forms of β-glucans are those comprising D-glucose units with (1,3) links and with side-chains of D-glucose attached at the (1,6) position.
B-glucan chemical
β-glucan sources in nature


One of the most common sources of β(1,3)D-glucan for supplement use is derived from the cell wall of baker’s yeast (Saccharomyces cerevisiae). However, β(1,3)(1,4)-glucans are also extracted from the bran of some grains, such as oats and barley, and to a much lesser degree in rye and wheat. The β(1,3)D-glucans from yeast are often insoluble. Those extracted from grains tend to be both soluble and insoluble. Other sources include some types of seaweed,and various species of mushrooms, such as reishi, shiitake, Chaga and maitake.
β-glucan and cholesterol
Several health claims requests were submitted to the EFSA NDA Panel (Dietetic Products, Nutrition and Allergies), related to the role of β-glucans in "maintenance of normal blood cholesterol concentrations and maintenance or achievement of a normal body weight". In July 2009, the Scientific Committee issued the following statements: On the basis of the data available, the Panel concludes that a cause-and-effect relationship has been established between the consumption of beta-glucans and the reduction of blood cholesterol concentrations.
A major criticism of these studies has been that people who consume large quantities of food such as oat bran may reduce their intake of foods containing saturated fatty acids. The lower cholesterol may therefore be a consequence of a low-fat diet rather than being attributable to the effect of the oat bran itself. This was the conclusion of a recent randomized, doubleblind study that attempted to maintain a constant amount of fat. In that study it was concluded that oat bran had little cholesterol-lowering effect and that high-fiber and low-fiber dietary grain supplements reduce serum cholesterol to about the same degree .
Evidence suggests that some soluble fibers bind bile acids or cholesterol during the intraluminal formation of micelles .The resulting reduction in the cholesterol content of liver cells leads to an up-regulation of the LDL receptors and thus increased clearance of LDL cholesterol. However, increased bile acid excretion may not be sufficient to account for the observed cholesterol reduction. Other suggested mechanisms include inhibition of hepatic fatty acid synthesis by products of fermentation (production of short-chain fatty acids such as acetate, butyrate, propionate); changes in intestinal motility; fibers with high viscosity causing slowed absorption of macronutrients, leading to increased insulin sensitivity; and increased satiety, leading to lower overall energy intake.

Am J Clin Nutr. 1999 Jan;69(1):30-42.
Cholesterol-lowering effects of dietary fiber: a meta-analysis.
Brown L, Rosner B, Willett WW, Sacks FM.
N Engl J Med. 1990 Jan 18;322(3):147-52.
Comparison of the effects of oat bran and low-fiber wheat on serum lipoprotein levels and blood pressure.
Swain JF, Rouse IL, Curley CB, Sacks FM.
Antitumor effects
The tumoricidal properties of β-glucans have been studied in several in vitro and in vivo animal models. In a mouse model study, β-1,3 glucan in conjunction with interferon gamma inhibited tumors and liver metastasis. In some studies, β-1,3 glucans enhanced the effects of chemotherapy. In a mouse carcinoma model, β-1,3 glucans did not reduce tumor incidence, but were associated with reduced mortality in combination with cyclophosphamide. In human patients with advanced gastric or colorectal cancer, the administration of β-1,3 glucans derived from shiitake mushrooms, in conjunction with chemotherapy, resulted in prolonged survival times.
Beta-glucans participate in the processes of repair, metabolism and detoxification, and affect the overall health of the body counteract the pathological conditions of reactive oxygen and nitrogen and the processes in which they participate. Reactive oxygen species (ROS) and nitrogen (RNS) play an important role in the pathogenesis of many diseases. Production of ROS is an integral part of aerobic metabolism of cells. Physiological concentrations of ROS play an important role in proper functioning of many cellular processes, and their overproduction occurs during induced oxidative stress. Very closely associated with oxidative stress is nitrosative stress. Nitric oxide (NO) reacts with molecular oxygen, superoxide anion radical and metal cations to give more reactive oxygen species. Reactive oxygen and nitrogen react with proteins to cause impairment of their function by oxidation or nitrosylation of amino acid residues, which can direct the path of apoptotic cells. In addition, nitric oxide enhances the effect induced by cyclooxygenase and becomes a mediator of inflammation.
The active components of yeast-derived β-glucan exert their unique immune stimulating functions by binding specifically to complement receptor 3 (CR3) via lectin-like domain (LLD) and activating CR3 to promote cellular cytotoxicity of iC3b-coated cancer cells. In addition, particulate yeast-derived β-glucan stimulates both innate and adaptive anti-tumor immune responses.

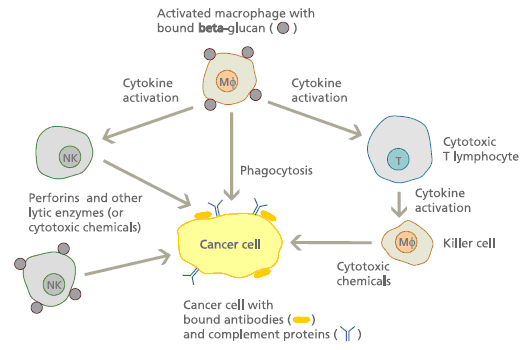
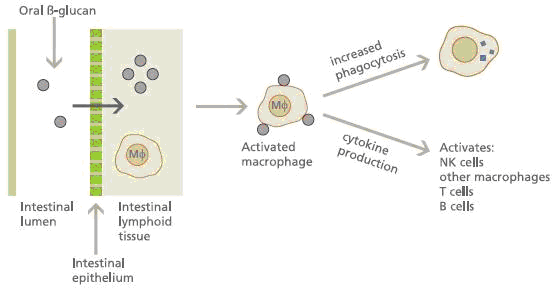
As an immunomodulating agent, β-glucan acts through the activation of innate immune cells such as macrophages, dendritic cells, granulocytes, and natural killer cells. This activation triggers the responses of adaptive immune cells such as CD4+ or CD8+ T cells and B cells, resulting in the inhibition of tumor growth and metastasis. Reports have shown that β-glucans exert multiple effects on cancer cells and cancer prevention.
Furthermore, in recent research has emerged that introduction of a sulfate group on the single chain lentinan increased antitumor activity due to increased binding of the β-glucan to receptors on immune cells via electrostatic interactions that activated the immune response., both the single and triple helices had strong TNF-a and IL-6 cytokine releasing activity in a macrophage cell line . In addition, a partially opened single helix induced more nitric oxide than did the original triple helix . Activation of innate immunity by β-glucan is initiated by its binding to the specific β-glucan receptor dectin-1 on macrophages and DCs or to CR3 on granulocytes and natural killer (NK) cells . β-Glucans also directly activate adaptive immune cells such as B cells , T cells, NK cells , eosinophils , and neutrophils .
Masuda et al. showed that highly purified, soluble β(1,3)(1,6)-glucan obtained from Grifola frondosa rapidly induced GM-CSF production through dectin-1-independent ERK and p38 MAPK activation. Subsequently, β-glucan induced GM-CSF-enhanced proliferation of resident macrophages and dectin-1 expression, which permitted dectin-1-mediated TNF-α induction through the Syk pathway .
Eur J Immunol. 2013 May;43(5):1220-30. doi: 10.1002/eji.201242841. Epub 2013 Mar 25.
β-Glucan enhances antitumor immune responses by regulating differentiation and function of monocytic myeloid-derived suppressor cells.
Tian J, Ma J, Ma K, Guo H, Baidoo SE, Zhang Y, Yan J, Lu L, Xu H, Wang S.
Med. Chem. giugno 2013; 13 (5) :699-708.
The Effects of β-glucans on Cancer MetastasisYoon TJ , Koppula S , Lee KH .
Curr Med Chem. 2012;19(25):4298-305.
Anti-tumor monoclonal antibodies in conjunction with β-glucans: a novel anti-cancer immunotherapy.
Xiang D, Sharma VR, Freter CE, Yan J.
Pol Merkur Lekarski. 2012 Oct;33(196):217-20.
[Beta-glucan as a natural anticancer agent].
Jurczyńska E, Saczko J, Kulbacka J, Kawa-Rygielska J, Błazewicz J.
b-glucan and immune system
β-Glucans,as already introduced previously, are known as "biological response modifiers" because of their ability to activate the immune system. Immunologists have discovered that receptors on the surface of innate immune cells called dectin-1 and complement receptor 3 (CR3 or CD11b/CD18) are responsible for binding to β-glucans, allowing the immune cells to recognize them as "non-self".
With respect to pattern recognition molecules in serum, complement proteins can recognize pathogens and subsequently be activated by the classical, alternative, or lectin pathways. All three culminate in activation of C3, the central step in complement activation, which is then followed in the final steps by formation of the cytolytic membrane attack complex (MAC). After the initial C3 proteolytic activation step, nascently activated C3b can covalently associate with the carbohydrates or proteins present on the surface of the pathogen. This initial process of C3b attachment to pathogens is followed by further inactivation of bound C3b by its proteolysis to iC3b, followed by further degradation to C3d. Complement opsonization of pathogens can lead to either direct killing of the pathogen by formation of MAC or recognition and destruction of the C3 fragment-opsonized pathogen by cell-associated complement receptors, CR1, CR2, CR3, or CR4 on leukocytes.
In addition to the pathways, signals from dectin-1 may follow another Syk-independent route. Dectin-1 could trigger a Raf1-mediated pathway that modulates NF-κB activation .
In this case, Raf-1 binds to the cytoplasmic domain of dectin-1 and increases p60 phosphorylation independent of Syk, which also occurs in the Syk-dependent pathway. This pathway exerts a synergistic effect along with β-glucan signaling. In addition, this pathway may serve as an alternative signal transduction pathway when the Syk-dependent pathway is blocked.
Another class of dectin receptor was recently discovered. This receptor, which was named as dectin-2, recognizes high mannose and α-mannan structures. Mannan is a polymer of mannose that is usually found in yeast, bacteria and plants. In contrast to dectin-1, dectin-2 lacks an ITAM motif in the cytoplasmic domain. Dectin-2 consequently employs the Fc receptor γ recep (FcRγ), which contains an ITAM motif . When FcRγ associates with dectin-2, Syk is recruited to the ITAM domain. The rest of the dectin-1 and dectin-2 signal transduction pathways are believed to be identical.
Currently, it is not known whether dectin-1 induces pathogen-specific adaptive immunity. However, dectin-1 contributes to the adaptive immune responses as well as autoimmune diseases and immune tolerance . Activated dectin-1 is involved in acquired immunity through enhanced expression of various cytokines, which subsequently promote the differentiation of Th17 cells. Dectin-1 may help modulate Th17 differentiation by promoting IL-6 and IL-23 by TLR2 along with concurrent inhibition of IL-12 by Th1. In addition, dectin-1 may play a role in acquired immunity by rapid antigen presentation through enhanced phagocytosis and ROS production.
In conclusion:

Immunol., 12 August 2013 10.3389/fimmu.2013.00230
Binding of soluble yeast β-glucan to human neutrophils and monocytes is complement-dependent.
Nandita Bose *, Anissa SH Chan , Faimola Guerrero, Carolyn M. Maristany, Xiaohong Qiu, Richard M. Biothera, Bloomington, MN, Stati Uniti d'America
Biomolecules and Therapeutics.
Immunomodulation of Fungal β-Glucan in Host Defense Signaling by Dectin-1.
The Korean Society of Applied Pharmacology 2012.20.5.433
Medicina (Kaunas). 2007;43(8):597-606.
Effects of beta-glucans on the immune system.
Akramiene D, Kondrotas A, Didziapetriene J, Kevelaitis E.